Claes_A
Medlem
-
Gick med
-
Senaste besök
All aktivitet
-
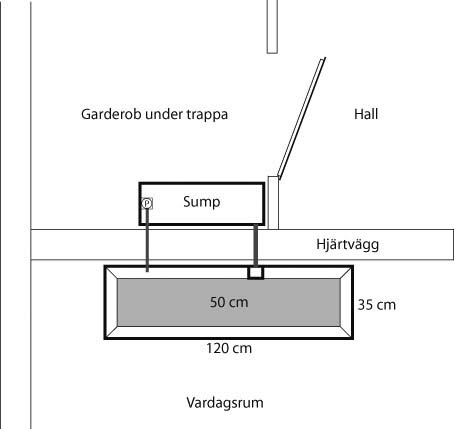
Efter några år i Tyskland har jag nu flyttat hem igen och funderar på att sätta upp ett nytt saltvattensakvarium. Därför startar jag denna planeringstråd så att jag kan bolla idéer och få mina idéer utvärderade. För tillfället finns det ingen helt klar plan vad akvariet skall innehålla och jag tvivlar lite på att jag har energin som krävs för att hantera de mest känsliga korallerna. Å andra sidan finns det en hel del begränsningar. Jag är uppväxt med saltvattensakvarier (arv från farsgubben) och har redan sovit bredvid brummande pumpar, heta metallhalogenlampor och förstört både parkettgolv och plastmattor genom översvämningar. Den tiden är förbi. Det nya akvariet skall vara knäpptyst (får inte störa ljudet från elden i kakelugnen), ljusföroreningar skall elimineras och värmefrågan måste vara löst (dvs inga onödiga mängder med elkonsumerande pumpar etc). Jag har hittat en solfri placering i vardagsrummet som erbjuder möjlighet att placera en sumpen i ett grannutrymme under husets trappa. Jag ser framför mig hur vattnet leds rakt igenom väggen till sumpen utan större fall. Genom att lägga sumpen i nivå med akvariet undviker jag därmed både ljud och elkonsumerande tryckpumpar. Vidare tror jag att detta minimerar risken för översvämningar - i alla fall om jag borrar två hål i akvariet och därmed får två redundanta utlopp. Idealt sett står cirkulationspumpen (pumparna) i sumpen för merparten av cirkulationen i akvariet. Ljudskapande utrustning (t ex skummare) placeras i sumpen och därmed inte i vardagsrummet. Här är en skiss på tänkt placering av akvarium och sump: Vattenreningsmässigt funderar jag på skummare med ozon (kanske t o m med träutströmmare för att minimera ljudet), DSB (väldigt tyst och energieffektivt) och eventuellt någon typ av biofilter som startas som ett vanligt nitrificerande biobollsfilter (Jag har tagit intryck av Lasse!) under de första veckorna för att sedan när akvariet är stabilt konverteras till ett denitrificerande filter med kolkällstillsats. Ljusmässigt funderade jag först på LED men jag förstår att T5 ger mer ljusenergi per insatt energi så jag lutar åt det. Metallhalogen bygger för mycket ovanför akvariet och ger för mycket värme- och ljusföroreningar. Tror ni mina krav på akvariet är realistiska?
-
Jag har kört mycket faxesten förr och visst ser de bra ut efter något år eller så. Men kalksten av typen faxe är väldigt kompakt och har knappast samma typ av hålrum som bra levande sten har. Så kanske ett bättre alternativ till artficiell levande sten är att använda den äkta varan istället. Förutom själva utmaningen i att lyckas tillverka artificiell levande sten så ser jag en potential att göra optimala och spännande inredningar i akvariet. Och om den tillverkade stenen dessutom har en porstruktur snarlik den äkta varan så desto bättre!
-
Jag har funderat lite på att tillverka artificiella levande stenar med cementbas. Tydligen är ett problem med artificiella levande stenar att de sällan har de fina håligheter som god autentisk levande sten har. En lösning som prövats för att få dessa håligheter är att tillsätta salt vid tillverkningsprocessen; saltet kan efter att cementen hårdnat tvättas bort vilket lämnar små hålrum. Problemet tycks dock vara att saltet försvagar cementen avsevärt så att stenarna lätt kan brytas sönder. Jag har funderat på lite andra komponenter för att tillverka artificiell levande sten. 1. Perlit som ballast Ett material som slår mig som idealt som ballast för stenen är perlit. Det är ett glasmineral som efter upphettning (expansion) bildar vita korn av sintrat glas. Porerna är öppna så att materialet lätt suger upp vatten. Perlit används av det skälet ofta som växtodlingssubstrat. Vad jag förstår skall Perlit vara ett ganska inert material som inte ger ifrån sig en massa metaller och elände. Ref: http://www.dicalite-europe.com/chemrock-perlite.htm Tror ni att perlit kan skapa cementsten med större andel håligheter än andra ballaster? 2. Socker istället för salt Salt känns som ett dåligt val för cementgjutning. Det är en jonisk förening som jag tror försvagar cementen avsevärt (någon som vet mer?). Istället har jag funderat på att använda vanligt socker. Det är inte joniskt och borde till skillnad mot salt inte alls reagera med cementen. Men i likhet med salt är det lätt att tvätta ur stenen med vanliga vattenbad. 3. Fosfat och snäckskalsfråga Jag förstår att snäckskal som ballast ser snyggt ut men introducerar fosfat. Är fosfaten bunden i själva skalet som mineral (kalciumfosfat t ex) eller kommer den från organiska föreningar som förorenar skalet? Dvs kan fosfaten tvättas ur snäckskalen om man avlägsnar de organiska föroreningarna (NaOH, klorin etc)?
-
Och en tråkig söndag ett halvår senare känner jag mig manad... Socker (låt oss välja glukos/druvsocker/dextrosol för enkelhetens skull) är en mycket snabb och bra energikälla för bakterier precis som för andra organismer (jämf människor). Om socker finns tillgängligt kommer bakterier att använda det som sin främsta energikälla (de tom slutar använda andra energikällor) och de kan använda det på två sätt: respiration och fermentering/jäsning. Om de respirerar det får de ut maximal energi ur det men processen går långsammare och syre går åt. Om de fermenterar det går processen snabbt men de får ut mindre energi per molekyl socker och det krävs inget syre. De flesta mikroorganismer kommer samtidigt att både fermentera och respirera tillgängligt socker. Socker kan respireras under syrefattiga förhållanden där nitrat används istället för syre och därmed ge upphov till denitrifikation. Etanol kan till skillnad mot socker inte fermenteras. Enda sättet att kräma ut energi från etanol är genom respiration (där denitrifikation är en variant!). Etanol är till skillnad mot socker heller inte ett populärt val bland alla organismer. Många bakterier, växter och djur har inte ens de rätta enzymerna (alkoholdehydrogenaser) för att kunna använda etanol som energikälla. Baserat på detta kan man tänka sig att tillsats av etanol och socker (glukos t ex) kan ge lite olika effekt på akvariet. Glukosen kan användas av alla möjliga organismer som energikälla och kommer med säkerhet att tas upp av akvariets samtliga invånare vare sig de denitrificerar eller ej. Etanolen kan inte användas av alla organismer och man kan tänka sig att den kanske speciellt gynnar bakteriepopulationer som denitrificerar (spekulation).
-
Tillbaka till trådens topic! Problemet är väl som alltid att vi till att vi inte ens är säkra på grundobservationen, dvs att närvaron av Aquaphosen skapar någon typ av alger/cyanobakterier som är bruna. Om vi nu ändå antar att detta stämmer (dvs kan reproduceras under kontrollerade former) kan vi ju fråga oss varför. Och egentligen har vi väl bara två klara förklaringsmodeller: 1. Aquaphosen släpper ut någonting som gynnar växten av dessa alger (järn, kisel, fosfat, etc) 2. Aquaphosen binder något vars brist hämmar åtgången av ett annat näringsämne som i sin tur gynnar algerna. Jag skulle sätta mina pengar på förklaringsmodell nr 1. Om det sedan är järn, kisel eller något annat vågar jag inte gissa. Troligen är det inte fosfat för jag tror inte att systemet får en ökad tillgång på fosfat bara för att man använder en fosfatbindande massa (om inte massan sett _väldigt_ höga nivåer fosfat under början av sin livstid och sedan läcker ut det). Sen är jag inte helt säker på om fosfat verkligen är tillväxtbegränsande (eller på annat sätt gynnar) tillväxt av dessa bruna alger. Förklaringsmodell 2 köper jag inte riktigt för den är krånglig och jag har svårt att tro att massan kan vara så effektiv att den helt sänker nivån av något näringsämne som är viktigt för säg koralltillväxten. Spårämnen lär det i alla fall inte röra sig om eftersom dessa per definition alltid finns i låga koncentrationer och därmed har svårt att helt nollas av bindningsmassor.
-
Vad vi fann för några veckor sedan i artikeln gällande Rowaphos var att jämvikten verkade vara långt förskjuten till bunden fosfat. Dvs massan var effektiv även vid ganska låga koncentrationer (helt i linje med vad en massa folk observerat hemma i sina akvarier). Vidare verkade bindingen ske snabbt vilket är vad man kan förvänta sig av jonbytarlika massor. Och här kommer min reaktion på citatet ovan: var jämviktförhållandet ligger (bunden fosfat eller fritt i vattenmassan) bestäms inte enbart av bindningshastigheten, utan av förhållandet (kvoten) bindningshastighet ("rate on") kontra frisläppningshastighet ("rate off"). Precis som Fredrik skriver ovan ger därför en hög bindningshastighet och låg frisläppningshastighet en massa med jämvikten långt åt bundet fosfat - och massan är dessutom snabb. Men även massor med en låg bindningshastighet kan ha jämvikten långt åt bunden fosfat förutsatt att frisläppningshastigheten är ännu sämre. En sådan massa skulle vara väldigt långsam.
-
Vad gäller proteiner lösta i vattnet: Eftersom det kräver en väldans massa energi för att bryta peptidbindingarna i proteinerna så måste nedbrytningen katalyseras av enzymer. Vi kan nog nästan utgå från att dessa enzymer kommer att produceras av bakterier eftersom de dominerar våra akvarier. Helt i linje med detta utsöndrar bakterier mängder med enzymer (av typen proteaser) som kan bryta ned proteiner till aminosyror och små peptider utanför cellen. Dessa importeras sedan in i cellen och försörjer bakterierna med främst kväve och energi.
-
För att aminosyrorna skall kunna vidhäfta en negativt laddad luftbubbla (eller en annan negativ partikel som lyfts ut via skummaren) måste den ha motsatt laddning - dvs positiv. Att bara vara polär hjälper inte. Arginin, histidin och lysin har alla en sidogrupp som är positivt ladded och kan därmed binda negativt laddade partiklar, men med tanke på den höga salthalten i vattnet torde det inte ske någon binding eftersom främst natriumjoner kommer att konkurrera ut deras bindning. Andra aminosyror är mer utpräglat amfipatiska - dvs de är laddade men har en oladdad fettlöslig sidokedja (tryptofan är ett bra exempel). Kanske kan dessa via sin fettlösliga sidokedja (som då kan tänkas binda nåt fettlösligt eller luftfasen) plockas ut av skummaren? Typiskt för dem är också att det är lite si och så med vattenlösligheten. Fast summa summarum tror jag inte fria aminosyror exporteras nämnvärt av skummaren. De tas nog upp av främst bakterier istället och sen exporteras bakterierna eller deras nedbrytningsprodukter (bla proteiner igen).
-
Det har nämnts i tråden att aminosyror skummas bort men jag ställer mig frågande till det. Aminosyror är små laddade molekyler och torde inte alls vara speciellt lämpad att bygga skum med vad jag förstår. Proteiner däremot innehåller oftast både laddade och hydrofoba (fettlösliga) delar och kan därmed placera sig så att den laddade sidan pekar mot vattenmolekylerna och de hydrofoba mot gasfasen. I alla fall på labbet har jag aldrig sett en benägenhet för aminosyralösningarna att bilda skum. Annat är det ju med proteinlösningar...
-
En fråga om man resonerar om aminosyror och koraller är givetvis om vissa aminosyror är essentiella överhuvudtaget. Normalt sett brukar man nämna 11 aminosyror som essentiella i djurvärlden: De tre besläktade grenkedjade aminosyrorna leucin, isoleucin, valin; de ringinnehållande fenylalanin, tryptofan och tyrosin; de positivt laddade histidin och lysin; de svavelinnehållande cystein och metionin samt threonin (stavas på svenska?). Vissa av dessa är helt essentiella för att djur saknar biosyntesvägar för dem (de grenkedjade är ett paradexempel) andra är delvis essentiella i olika delar av livet för att biosyntesen inte räcker till (cystein är ett krav för barn exempelvis). Då jag för några år sedan grävde i vad som utretts vad gäller koraller fann jag att koraller inte tycks likna andra djur vad gäller aminosyrakrav. I själva verket verkar threonin vara den enda aminosyra som inte kan syntetiseras av koraller(http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=1218179). Det verkar alltså som om korallerna till skillnad från andra djur behållit biosyntesvägarna för samtliga normala aminosyror utom threonin och har därmed inte så många essentiella aminosyrokrav. Jag skall gräva lite djupare i det hela då jag får tid.
-
Det stämmer att det finns en fördel att importera aminosyror för en cell även om de inte är essentiella. Argumenten är tre: 1. (Som sagt) Det kostar energi att syntetisera aminosyror. 2. Aminosyror är den huvudsakliga kvävekällan för alla celler. Cellen upprätthåller en jämn nivå av aminosyrorna glutamin och glutamat (smakförstärkaren ni vet!) och i princip alla andra kväverika ämnen i cellen syntetiseras med start från dessa två aminosyror. I princip är det denna reaktion som sker (kan katalyseras att gå åt båda hållen av olika enzymer): glutamat + ammonium <=> glutamin 3. Aminosyror kan användas för att försörja cellen på energi.
-
Lustigt nog råkar jag ha doktorerat på cellers aminosyraupptag och vissa saker står klart för mig: Fria aminosyror i vattenmassan utgör troligen inte en naturlig diet för koraller pga att dessa molekyler är rena hårdvaran som kvävekälla för mikroorganismer. Dvs de kommer att konsumeras omgående och koncentrationerna hålls därmed låga i vattnet kring korallreven. Däremot konsumerar ju koraller aminosyror som de får genom nedbrytning av plankton och annan fångst i maghålan. Denna nedbrytning av proteiner till aminosyror och små peptider (vanligen di- och tripeptider) är ett grundkrav för att kunna importera molekylerna in i cellen genom plasmamembranet - proteiner är på tok för stora för att importeras. Själva transporten över plasma membranet sköts av transportörproteiner som ofta kallas permeaser och som har olika substratspecificitet. Dvs en typisk cell har ett ganska stort antal permeaser i plasmamembranet som förser cellen med olika typer av näringsämnen. De flesta celler har ett helt batteri av just aminosyrapermeaser som tar upp olika typer av aminosyror eftersom aminosyror är energimässigt kostsamma att syntetisera och utgör den klart bästa kvävekällan för cellen. Och här kommer vi till essensen i resonemanget: koraller borde kunna ta upp fria aminosyror från vattenmassan utan problem om det doseras till akvariets vattenmassa genom sina permeaser. Frågan är bara om dessa aminosyror klarar sig tillräckligt länge i akvariets vattenvolym med tanke på att varenda mikroorganism (och alg!) också kommer att försöka ta upp fria aminosyror som tillsätts. Kanske leder aminosyratillsatsen bara till ökad baterietillväxt i akvariet som i sin tur blir foder till korallerna?
-
Randy-Holmes Farley har gjort en serie enkla och lättolkade experiment med kalciumhydroxid och kalkvatten: http://www.reefkeeping.com/issues/2005-01/rhf/index.php Enligt mitt tycke är artikeln ett måste att läsa om och om igen varje gång oklarheter dyker upp vad gäller kalkvatten, dess hållbarhet och hur mättnad nås. Detta mytdödarpapper visar att kemisk jämvikt (=mättnad) är omedelbar när kalciumhydroxiden blandas med vatten och att mättnad kan upprätthållas enkelt och billigt genom att bara tillsätta så pass mycket kalciumhydroxidpuder att det finns en mänd kvar på bottnen. Vidare ifrågasätter han kraftigt om en kalkvattensmixer fyller någon som helst arbetsbesparande funktion i och med att jämvikt nås så lätt och kalkvattnet är ganska stabilt över tiden.
-
Jag har nu slutligen läst igenom SIR-600-artikeln och har svårigheter att fastställa om experimenten verkligen ger några vettiga svar. Författarna rapporterar nämligen experiment där så mycket som 30 g SIR-600 används på så lite som 100 ml (1 dl!) vatten. Eftersom tillverkaren anger att materialet binder 1.3 mol monokatjonsekvivalenter per liter (motsvarar exempelvis 50 g kaliumjoner) torde 30 g av materialet med lätthet hunna binda större delen av kaliumet i havsvattnet i experimentet (100 ml havsvatten innehåller 1 mmol = 0.04 g kaliumjoner). Vidare anger tillverkaren att affiniteten för kaliumjoner är högre än för ammonium vilket skulle betyda att materialet i princip kan strippa den lilla vattenvolymen som används på kalium och endast kvarvarande kapacitet skulle då binda ammonium. Denna tolkning verkar också stämma eftersom att författarna anger att när mindre mängder zeolith användes (5 g) fick de sämre avlägsnande av ammonium. Jag hade hoppats att artikeln dokumenterade i alla fall en zeolith som kan binda ammonium med hög affinitet i saltvatten, men det verkar inte så. I avsaknad a "proof of principle" kvarstår endast tillverkarens uppgift att ZEOvit är en blandning av zeolither som faktiskt kan binda ammonium i saltvatten. Vad gäller tillgänglighet av eventuellt bundet ammonium i ZEOvit så finns vissa faktum att ta hänsyn till: - Bakterier (liksom alla celler) importerar ammonium via transportörer i plasma membranet. Sådana transportörer kan bara binda ammoniumjoner i lösning. Bundet ammonium är inte direkt tillgängligt. - Jonbytare står alltid i jämvikt med omgivande vatten och upprättar den jämvikten snabbt. Hur långt den jämvikten är driven till bunden eller fri jon beror helt på affiniteten. Om ZEOvit kan binda ammonium i saltvatten måste den ha en mycket högre affinitet för ammonium än andra vanliga joner som natrium och kalcium. Notera dock att i ett ganska komplext jonsystem som saltvatten kommer olika joner att tävla om bindningsplatserna och jonerna kommer därmed att mycket snabbt hoppa av och på materialet. Ammonium skulle därmed lokalt kunna vara tillgängliga för bakterier om affiniteten för ammonium och övriga joner var rätt avpassad. Om den sedan är det vad gäller ZEOvit eller om det ens är praktiskt möjligt vågar jag inte yttra mig om. (Jag börjar bli lite frusterad av bristen på fakta och funderar faktiskt på att köpa mig lite ZEOvit och testa om det ens binder ammonium i saltvatten...)
-
Notera att SIR-600 är kaliumselektiv och därmed kan kalium konkurrera med ammonium vad gäller bindning till mediet. Dvs ammonium kommer konstant att bytas ut mot kalium och sedan snabbt återbindas igen. Detta gör att ammonium inte är konstant bundet utan kommer att finnas "tillfälligt" i lösning i omedelbar närhet av mediet och därmed vara tillgängligt för bakteriernas importmaskineri.
-
Även Heidelberg prenumererar på blaskan ifråga så jag har också pdfn för Burgess et al. Släng mig ett email med en epostadress så kan jag skcka över filen om så önskas. Ska läsa den ikväll.
-
Lasse Den massiva expansionen av antalet vetenskapliga tidskrifter på senare år har gjort att universiteten spararat in på prenumerationer av specialtidskrifter som det ofta handlar om i vattenreningssvängen. Jag ska se vad Universität Heidelberg har att erbjuda vad gäller pdfen för artikeln på jobbet imorgon... Jag läser Kallmeyer på detta sätt: 1. Katjonbytare fungerar dåligt i saltvatten pga den höga salthalten men moderna zeoliter (e g ZEOvit) erbjuder en affinititet som är specifik för ammoniumjoner (en sterisk passning som en nyckel och ett lås) som gör att de fungerar ok även i närvaro av höga koncentrationer av andra katjoner. Andra joner kan helt enkelt inte konkurrera ut de elektrostatiska bindingarna som ammonium bildar med materialet eftersom de inte passar in. 2. Affiniteten för ammonium som finns hos ZEOvit skapar en lokalt högre koncentration av ammonium i den omedelbara närheten av materialet (i hålrummen) över tiden. Denna lokalt högre koncentration gör att systemets kvävebegränsade bakterier kan växa där. Dessa två viktiga punkter baseras givetvis på mitt försök till rationalisering (svengelska?) av hans populärvetenskapliga text. För att 2 skall vara sann måste ju 1 vara sann säger logiken i det här fallet så uppmätt ammoniumbindning i saltvatten vore intressant att se. Jag letade lite i databaserna igår kväll men hittade inte mer än den refererade artikelns abstract. Alla andra artiklar jag hittade betonar frånvaron av ammoniumbindning i saltvatten.
-
Och notera att nästan alla bakterier är ammoniumälskande i en situation av kvävebegräsning (vilken man kan tänka sig kan uppstå vid koltillsats och användandet av nån typ av massa som har god affinitet för ammonium). Inte bara nitrifierarna. Fast det vore ju kul att se lite experiment som påvisar ammoniumbindning i saltvatten av ZEOvitmassan...
-
Om ni inte har läst det förut så tänkte jag bidra med Jens Kallmeyer ritar och berättar om zeoliter i saltvatten: http://www.wetwebmedia.com/ca/cav1i3/zeovit/Zeolite_Filters/Zeolite_Filters.htm Jag tycker mycket av det han skriver låter rimligt men kanske underskattar han de heterotrofa bakteriernas funktion som näringsupptagare och överskattar de autotrofa nitrifierarna. Och hela resonemanget bygger ju på att man har tillgång till en zeolit med sådan specificitet för ammoniumjoner att den fungerar även vid höga salthalter. Jag har hittat stöd för att sådana finns i följande artikels abstract: http://www.springerlink.com/content/r402qj3qb9gwub04/
-
-
Som vanligt nådde ironin inte fram. Gud är han inte men det är mycket roande att läsa hans predikan. Och trots hans övertro på att han har funnit den "den rätta vägen" så tar han ju upp ett och annat guldkorn emellanåt.
-
Lästips! Nu hittade jag Bornemans svavelosande sågning av vodkatillsatser a la Mrutzek och Kokott igen. Kul läsning och han tar upp ett antal perspektiv som inte berörts i den här tråden ännu. Vidare får man lära sig att sandbäddar är den enda vettiga metoden för ett fungerande korallrevsakvarium: http://www.reefkeeping.com/issues/2004-11/eb/feature/index.php Tillbaka till Marine planted: Inspirerad av Bornesmans beskrivningar av bottenekologin runt reven och hur dessa bottnar omöjligen skulle kunna vara kolbegränsade utan snarare kvävebegränsande börjar jag fundera på om inte detta är värt att tänka på då man startar en sandbädd. Om vi utgår från att man startar med rent korallgrus i sitta akvarium så det det ju lång tid innan sandbädden kommer att börja fungera (jag tror det tog närmare ett år innan jag fick ordentlig reduktion i mitt senaste akvarium och då jag la ned det efter många år kunde jag konstatera att det fanns mängder av reducerande zoner i bottnen). Inympning av bakterier torde hjälpa akvariet att komma igång med nitrifikationen (som nog ändå inte sker annat än på ytan av sandbädden) men reduktiv nedbrytning av organiska ämnen och denitrifikation torde ta längre tid att etablera. Sandbädden är helt enkelt för ren och troligen kolbegränsad till en början. Så borde man inte tillföra en långsam kolkälla till sandbädden under det första halvåret eller så (sen blir ju sandbädden självfungerande). Rent praktiskt funderar jag på en jordbotten - kanske någon näve ordentligt urlakad och välkomposterad organisk jord (vi vill ju inte tillföra kväve och fosfor) i bottnen på sandbädden. Idén låter skrämmande även för mig!
-
Då kan jag passa på och fråga hur forumet (Stig?) ställer sig till faxekross kontra korallsand. Är de jämförbara och finns det en risk att introducera en oönskad fosfatkälla i något av fallen? Vidare, vad sägs om karbonatbaserad sand i bottnen och kiselbaserat dito ovanför?
-
Jag har under dagen roat mig med att fördjupa mina kunskaper i bakterieekologi i olika vattensystem. Alla artiklar rapporteras i princip samma typ av experiment: Lite vatten eller filtrat tas (från sediment eller från vattenpelaren) och ges tillsatser (kolkällor, kväve fosfor, etc) och sen får bakterierna tid att växa och kan man fastställa om bakteriernas tillväxt är begränsad an något ämne. Precis som Lasse och jag har framfört så rapporteras organiskt kol ofta vara begränsande för tillväxten av heterotrofa bakterier både i sjöar (sötvatten) och marina miljöer. Fosfor är också ofta begränsande, ibland tillsammans med kol. Sen det här med fritt svävande bakterier (bakterieplankton). Alla bakterier som används i artiklarna växer bra som bakterieplankton och kemostater (en apparat där bakteriekulturer i lösning kan hållas i en helt konstant miljö genom utspädning) är standardmetoden för att dra slutsatser om deras beteende. Noteras skall att tom litoautotrofer som Nitrosomonas, Nitrospira och Nitrobacter växer glatt i lösning på lab. Konsistent med detta kan man också hitta mängder med bakterier i den fria vattnenmassan som bakterieplankton i naturliga vatten. Det är alltså bakterier som inte växer på några som helst partiklar - precis som på lab. Men populationerna är kraftigt begränsade i sin tillväxt i brist på näring (kol eller fosfor igen!) och temperatur där det är kallare, så populationerna blir aldrig rejäla stora. Helt i linje med Lasses resonemang är partiklar väldigt viktiga som växtsubstrat för bakterier i vattensystem. Men anledningen är inte på något sätt att bakterierna behöver en yta att växa på utan att partiklarna utgör eller har näring associerat till sig. En artikel rapporterar tio gånger mer högre koncentrationer av begränsande näring (kol) i marint sediment jämfört med ovanliggande vattenpelare och pga detta var mängden heterotrofa bakterier i sedimentet enormt mycket större där jämfört med mängden heterotrofa bakterieplanktion. Men, då författarna tittade på deras nedbrytningskapacitet per cell (glukoskonsumtion) fanns ingen skillnad mellan bakterierna som växte på sedimentet och bakterieplankton. Slutsatsen jag drar är att heterotrofa bakterieplankton växer lika bra som bakterier på substrat men eftersom de oftast är näringsbegränsade i jämförelse med sin fastsittande kusiner så är de inte lika viktiga i många habitat (inklusive akvarier). Jag står därför envist fast vid att tillsats av en energrik kolkälla kan (och borde) leda till en blomming av heterotrofa bakerieplankton i akvariet om kol är begränsande för tillväxt. Disighet är helt i linje med bakterier som växer i lösning; standardmetoden för att mäta bakteriekoncentrationen med en spektrofotometer är just disighet (light scattering 600 nm) och mina kulturer blir bra grumliga då bakterierna växer. Sen blir begreppsförvirringen ganska rejäl då man talar om vilka föreningar som är i lösning och inte. Vissa ekologer verkar helt nöjda med att filtrera vattnet genom ganska grova filter för att ta bort partiklar. Det som går igenom filtret menar de är i lösning ("dissolved"). Dvs mindre partiklar som en kemist skulle säga varande i suspension kan en ekolog kalla för lösta. Det hela blir ganska subjektivt och rörigt. En annan källa till begreppsförvirring är att artiklarna om reningsverk ofta jobbar med kontinuerliga system. Dvs de spolar vatten genom sina filter och kollar vad som händer på vägen. Om bakterierna inte växer på filtret som en biofilm har de endast en mycket liten chans att påverka systemet eftersom att de delar sig långsammare än flödeshastigheten kräver - de spolas ut med badvattnet. Detta gäller ju inte i akvarier utan effektiva UV-filter. Det hela slutar i svensk konsensus: Det står helt klart för mig att biofilmer och sediment är de huvudsakliga bakteriehabitaten i akvatiska system som våra akvarier!
-
Lasse- det här med begränsningar av näring vid tillväxt. Det jag inte begriper ditt resonemang är vad du anser vara begränsande för bakterietillväxten i olika akvariesammanhang. Skulle ingen begränsning finnas så skulle bakterierna göra akvariet till en tjock bakteriesoppa. Det är precis vad jag skapar på lab varje dag genom att odla bakterier (i princip bara E. coli) i mycket rikt näringsmedium. Våra flaskkulturer stannar av i växten efter något dygn av tillväxt pga av magnesiumbegränsning eller energibegränsning (peptider och aminosyror fungerar som kol och energikälla). Varför växer inte alla möjliga heterotrofer till i akvariet? Vad är det som de saknar? Jag tror att de saknar en liten fördelaktig energikälla av kolbas som exempelvis socker i första hand och i andra hand en fördelaktig kvävekälla som aminosyror/ammonium. Dvs jag är ganska övertygad om att tillsats av 0,2% (w/v) glukos till akvariet kommer att skapa en stinkande grumlig bakteriesoppa som säkert då är både kväve och syrebegränsad.